Topographische Anatomie: Anatomische Propädeutik: Grundlagen der Embryologie
Wanderung der Urkeimzellen
BearbeitenIn der vierten Woche wandern die Urkeimzellen aus dem Dottersack in die Gonadenanlage; im Zölomepithel vermehren sie sich und differenzieren sie sich. Von jetzt an läuft die Differenzierung der Gameten geschlechtsspezifisch ab.
Entwicklung der Keimzellen
BearbeitenOogenese
BearbeitenDie Eizelle entsteht aus Urkeimzellen, die sich teilen und charakteristische Phasen mit typischen morphologischen Veränderungen durchlaufen. Sie erhalten beispielsweise ein Epithel, das aber nicht nur als Eizell-Schutz dient, sondern auch Hormone produziert, die vor allem auf das Epithel des Uterus einwirken.
Die Urkeimzellen teilen sich mitotisch, so dass im 5. Monat etwa 7 Millionen Zellen vorliegen. Diese Zellen werden "Oogonien" genannt und beginnen die Reifeteilung I, die sie bis zum Diktiotän fortführen. In diesem Stadium wird die Meiose aufgrund inhibierender Substanzen des Follikelepithels gestoppt, die Zellen heißen nun "Oocyten I" und sind diploid mit zwei Chromatiden pro Chromosom; ihre Anzahl ist währenddessen auf etwa eine Million gesunken. Die Oocyten I verharren in diesem Zustand vom 7. Entwicklungmonat bis zum Eintritt der Pubertät, wobei ein Großteil der Zellen untergeht. Mit dem Beginn der Pubertät sind nur mehr etwa 40.000 Zellen übrig (von denen schließlich nur etwa 1% zu Eizellen heranreifen). Weil sie ein flaches einschichtiges Epithel haben, werden sie morphologischen Kriterien folgend als "Primordialfollikel" bezeichnet.

Alle 28 Tage werden etwa 50 Oocyten I aktiviert, d. h. sie führen die Meiose fort und ändern zugleich ihre Gestalt: Die Meisoe setzen sie fort, indem sie
- die Reifeteilung I vollenden (sie sind somit diploid und enthalten je ein Polkörperchen, d. h. eine Cytosol-arme Nebenzelle, die im Zuge der Meiose I entsteht) und
- in die Reifeteilung II eintreten, und zwar bis zur Metaphase.
Zugleich ändern sie ihre Gestalt von Primär- zu Sekundär- und Tertiärfollikeln:
- Primärfollikel haben ein höheres einschichtiges Epithel als Primordialfollikel
- Sekundärfollikel besitzen ein mehrschichtiges Epithel (zwei oder mehr Schichten), dessen Zellen nun "Granulosazellen" genannt werden; zwischen Plasmamembran und Epithel befindet sich die Zona pellucida (von der Eizelle produzierte Glykoproteinhülle), außen schließt sich dem Epithel die Theka folliculi an; durch die sekretorische Tätigkeit der Granulosazellen entstehen Löcher innerhalb des Epithels, die sich zum Antrum folliculare vereinigen.
- Bei den Tertiärfollikel sind die eigentliche Zelle, die umgebende Zona pellucida sowie die umgebenden Follikelepithelzellen (= Corona radiata) an den Rand verdrängt und erheben sich dort als Cumulus oophorus (Eihügel) ins Antrum; die umliegende Theka folliculi differenziert sich in eine epithelähnliche Theka interna (stellt Androgene her, die von den Granulosazellen zu Östrogenen aromatisiert werden) und eine bindegewebige Theka externa.
Durch die Wirkung von FSH aus der Hypophyse wird ein Tertiärfollikel selektioniert und vergrößert sich zum sprungreifen Graaf-Follikel.
Hormonale Veränderungen (LH-Peak) bewirken, dass sich der Eihügel ablöst und der Graaf-Follikel aufplatzt. Der Eihügel wird von den Fimbrien aufgefangen und gelangt auf diese Weise in die Tuba uterina. Der übriggebliebene Graaf-Follikel füllt sich mit Blut (Corpus rubrum) und verfärbt sich dann gelblich (Corpus luteum), weil die Granulosa- und Thekazellen Hormone produzieren (u. a. Progesteron, das für die Aufrechterhaltung des Uterusepithels (Endometrium) nötig ist). Nun kann eine Befruchtung stattfinden oder ausbleiben.
- Wenn keine Befruchtung stattfindet, degeneriert das Corpus luteum (in diesem Fall auch als "Corpus menstruationis" bezeichnet) zum Corpus albicans und geht schließlich ganz zugrunde; weil dadurch auch die gerade erwähnte Hormonproduktion zum Stillstand kommt, löst sich das Endometrium ab, die Monatsblutung (Mens) tritt ein.
- Findet dagegen eine Befruchtung statt, dann bleibt das Corpus luteum (in diesem Fall auch als "Corpus graviditatis" bezeichnet) bestehen, und zwar aufgrund von Rückwirkungen des sich einnistenden Eizell-Derivats (Blastocyste); das Corpus luteum vergrößert sich sogar und hält dadurch die Hormonproduktion aufrecht.
|
|
Merke Die Zahl der weiblichen Urkeimzellen schrumpft von 7 Millionen während der Embryonalperiode auf 40.000 zu Beginn der Pubertät. Während dieser Zeit befinden sie sich im Diktiotän, sie pausieren also in der Prophase der Meiose I. Die beim Zyklusbeginn durch hypophysiäre Hormone selektionierte Eizelle führt die Meiose bis zur Metaphase der Meiose II fort und verharrt in dieser Phase bis zum Eintreffen eines Spermiums. Primärfollikel haben ein flaches Epithel; Tertiärfollikel besitzen eine komplexe Hülle aus Zona pellucida, Granulosazellen (mit Antrum), Theka interna und Theka externa. Zwischen Corpus luteum und Blastocyste existiert eine positive Feedbackschleife: hCG aus dem Syncytiotrophoblasten der Blastocyste fördert das Corpus luteum, Progesteron aus dem Corpus luteum fördert das Endometrium und somit auch die Blastocyste. Wenn keine Befruchtung stattgefunden hat und folglich keine Blastocyste vorliegt, dann gibt es auch kein Feedback - das Corpus luteum geht zugrunde und somit auch das Endometrium (Mens wegen Progesteron-Entzugs). |
|---|

Spermatogenese
Bearbeiten

Die Spermatogenese findet im Hoden statt, und zwar in der Wand von kleinen Kanälchen (Tubuli seminiferi). Sie beginnt mit den A-Spermatogonien und teilt sich in drei Phasen auf: eine Vermehrungsphase, eine Reifungsphase und eine Differenzierungsphase (setzt erst in der Pubertät ein). Mit jeder Phase wandern die Zellen ein Stück weiter in Richtung Kanälchen-Lumen.
- Vermehrungsphase: A-Spermatogonien vermehren sich mitotisch, d. h. es entstehen zwei Spermatogonien.
- Die eine dient als Reserve (A-Spermatogonie),
- die andere (B-Spermatogonie) tritt in die Reifungsphase ein.
- Reifungsphase
- Die B-Spermatogonie verdoppelt ihre DNA (S-Phase) und heißt dann "Spermatocyten I".
- Die Spermatocyte I vollzieht die Reifeteilung I, woraus zwei Zellen hervorgehen (Spermatocyten II), die über Interzellularbrücken miteinander in Kontakt bleiben.
- Die zwei Spermatocyten II vollziehen die Reifeteilung II, es entstehen schließlich vier Spermatiden.
- Differenzierungsphase (erst mit Eintritt in die Pubertät): Die Spermatiden ändern ihre Gestalt und verlassen dann die Kanälchen-Wand; sie heißen nun "Spermatozoen" und haben einen charakteristischen Aufbau aus Kopf, Hals und Schwanz:
- Der 4 µm große Kopf besteht aus
- Akrosom (ein Lysosomen-Derivat, das lytische Enzyme enthält)
- Kern (stark kondensiert),
- außerdem befinden sich am Kopf Riechrezeptoren.
- Der Hals enthält
- Zwei Zentriolen
- Basalplatte, über die das Axonema des Schwanzes an den Kopf andockt
- Der Schwanz teilt sich in Mittelstück, Hauptstück und Endstück. Hauptbestandteil ist das Axonema (Geißel, 9x2+2-Aufbau), das von Ringfasern umgeben ist. Im Mittelstück liegen zusätzlich Mitochondrien vor, die Energie in Form von ATP für den Geißelschlag liefern.
- Der 4 µm große Kopf besteht aus
|
|
Merke Undifferenzierte männliche Urkeimzellen liegen einzeln im peripheren Bereichen der Tubuli seminiferi, ausdifferenzierte Spermien befinden sich dagegen am Lumen. Sich differenzierende Urkeimzellen sind über Interzellularbrücken miteinander verbunden. Ein Spermium besteht aus Kopf (mit Kern und Akrosom), Hals (mit Zentriolen und Basalplatte) und Schwanz (mit Mitochondrien und Axonema). |
|---|
Fertilisation
Bearbeiten
In einem Ejakulat schwimmen etwa 300 Millionen Spermien. Nachdem sie verschiedene Barrieren überwunden haben (sauerer pH in der Vagina, Schleimpfropf im Uterushals, Engstellen am Tubeneingang), gelangen sie in die Tuba uterina und durchlaufen dort einen Reifungsprozess (Kapazitation), wobei sie ihre Oberflächeneigenschaften verändern. Beim Kontakt mit der Eizelle in der Tuba uterina kommt es zur Akrosomenreaktion, d. h. die im Akrosom gespeicherte Enzyme (Hyaluronidase und proteolytische Enzyme) werden frei, und der Kopf durchdringt zuerst die Corona radiata und daut anschließend die Zona pellucida der Eizelle an. Hierauf verschmelzen die Membranen von Spermium und Eizelle miteinander, der Kern des Spermiums (= männlicher Vorkern) tritt in die Eizelle ein, während Hals und Schwanz des Spermiums abgekoppelt werden. Dies leitet die Eizellreaktion ein:
- Die Eizelle wird depolarisiert und dadurch aktiviert
- Der Inhalt kortikaler Granula wird freigesetzt, was die Struktur der Zona pellucida verändert
Beides verhindert, dass weitere Spermien in die Eizelle eindringen können (Polyspermieblock).
Anschließend vollendet die Eizelle die Meiose II, so dass der weibliche Vorkern entsteht. Männlicher und weiblicher Vorkern replizieren ihre DNA, anschließend verschmelzen sie miteinander (= Befruchtung, Fertilisation; allerdings wird hierbei keine Kernhülle gebildet); auf diese Weise ist – im Allgemeinen etwa 24 Stunden nach der Ovulation – eine Zygote entstanden. In den folgenden ca. 38 Wochen wird sich der Keim bis zum geburtsreifen Fetus weiterentwickeln (gerechnet wird ab der letzten Menstruation, d. h. 38 + 2 Wochen = 40 Wochen).
|
|
Merke Der Ablauf der Fertilisation lässt sich folgendermaßen gliedern: Kapazitation des Spermiums - Akrosomenreaktion des Spermiums mit Durchdringen von Corona radiata und Zona pellucida - Membranverschmelzung mit Freisetzung des männlichen Vorkerns in die Eizelle - Eizellreaktion - Entstehung des weiblichen Vorkerns - Verschmelzung von männlichem und weiblichem Vorkern. |
|---|
Implantation
BearbeitenIn der Implantationsphase setzt sich der Keim im Endometrium fest. Man unterscheidet eine Präimplantationsphase, in der die wesentliche Differenzierung in Embryo- und Trophoblast erfolgt, und die Implantationsphase, in der sich der Keim in die Uteruswand einnistet.
Präimplantationsphase
Bearbeiten
Die Zygote vollzieht nun eine Reihe von Furchungsteilungen, wobei die Gesamtgröße des dabei entstehenden Zellverbands aufgrund der starren Zona pellucida konstant bleibt:
- Die erste Furchungsteilung erfolgt 30 Stunden nach der Fertilisation, es liegen dann 2 Blastomeren vor (2-Zell-Stadium).
- Über weitere Furchungsteilungen tritt die Eizelle zunächst ins 4-Zell-, dann ins 8-Zell- und schließlich ins 16-Zell-Stadium (Morula-Stadium, 3. Tag) und in höhere Zell-Stadien (32-Zell, 64-Zell-Stadium etc.) mit entsprechender Zahl von Blastomeren über; die Furchungen können aber auch asynchron verlaufen. Die Furchungsteilungen gewährleisten somit, dass die unausgewogene Kern-Plasma-Relation der Zygote (großes Cytoplasma, dazu vergleichsweise kleiner Kern) in ein ausgewogeneres Verhältnis überführt wird.
Während des 8-Zell- oder 16-Zell-Stadiums findet die Kompaktierung statt: die Zellen differenzieren sich in
- eine äußere Zellmasse (liegt der Zona pellucida an, die Zellen sind über wasserundurchlässige Tight Junctions miteinander verbunden) und
- eine innere Zellmasse.

Anschließend entsteht die Blastocyste:
- Die Zellen der äußeren Zellmasse fangen nun an, Wasser ins Innere der Morula zu pumpen (sie pumpen eigentlich Natrium sekundär aktiv, Wasser folgt dann passiv nach). Dadurch erweitern sich die Interzellularräume zwischen den Zellen der inneren Zellmasse und fließen zur Blastocystenhöhle zusammen. Infolgedessen werden die Zellen der inneren Zellmasse an einen Pol verdrängt. Somit lassen sich zwei Anteile unterscheiden:
- die innere Zellschicht: Embryoblast, aus dem später der kindliche Organsimus hervorgeht,
- die umgebende Hüllschicht der äußeren Zellmasse: Trophoblast, aus dem später die Plazenta hervorgeht.
- Zur selben Zeit verliert die Zellmasse die sie umgebende Zona pellucida und vergrößert sich dann, weil sie Flüssigkeit aus der Umgebung aufnimmt.
Diese vergrößerte, aus Trophoblast und Embryoblast bestehende Zellansammlung wird "Blastocyste" genannt.
|
|
Merke Die Zygote vollzieht eine Reihe von Furchungsteilungen, wobei die dabei entstehenden Zellen bis zum 8-Zell- oder 16-Zell-Stadium mehr oder weniger gleichwertig sind. Ab dem 8- oder 16-Zell-Stadium geht diese Gleichwertigkeit verloren, weil die äußeren Zellen Ernährungsfunktionen übernehmen und die inneren Zellen sich in den Dienst der Organismus-Entwicklung stellen. |
|---|
Implantationsphase (Nidation)
Bearbeiten
Die Blastocyste heftet sich mit Hilfe von Adhäsionsmolekülen und unterstützt durch lytische Enzymen und Signalstoffe, die eine mütterliche Immunantwort unterdrücken, an das Endometrium an, und zwar meist an der oberen Hinterwand des Uterus, seltener an der oberen Vorderwand (pathologische Einnistungsorte: Ovar, Tuba uterina, Uterushals, Bauchhöhle). Der Endometriums-nahe Teil des Trophoblasten wandelt sich daraufhin in ein Syncytium (vielkernige Riesenzelle) um und frisst sich – angetrieben von Hypoxie – mit Hilfe von lytischen Enzymen immer weiter in das Endometrium hinein, wobei sich immer mehr Zellen des Trophoblasten dem Syncytiotrophoblasten anschließen. Da der Syncytiotrophoblast bei seinem invasiven Vordringen Zellen des Endometriums zerstört, werden Nährstoffe (Lipide, Glykogen etc.) freigesetzt, die dem Syncytiotrophoblasten seinerseits als Nährstoffe dienen. Man unterscheidet nun zwei Anteile des Trophoblasten:
- den außen gelegenen Syncytiotrophoblasten, der auch die Aufgabe hat, Hormone (hCG) zu produzieren, und
- den innen gelegenen Cytotrophoblasten, der gewissermaßen als Nachschub für den Syncytiotrophoblasten dient; seine Reservezell-Kapazität ist jedoch am Ende der Schwangerschaft erschöpft, so dass der Cytotrophoblast zugrunde geht.
Schließlich ist die Blastocyste vollständig in das Endometrium eingedrungen (interstitielle Implantation, 8. Tag). Der Syncytiotrophoblast breitet sich dabei weiter invasiv aus, wobei er mütterliche Gefäße arrodiert, so dass mütterliches Blut in Lakunen innerhalb des Syncytiotrophoblasten einfließen kann; somit ist die Versorgung des Embryos sichergestellt.
|
|
Merke Die Blastocyste heftet sich zunächst an die obere Hinter- oder Vorderwand des Uterus an. Anschließend frisst sie sich mit Hilfe des Syncytiotrophoblasten (der sich aus Zellen des Cytotrophoblasten rekrutiert) in das Endometrium hinein; dabei arrodiert sie mütterliche Blutgefäße. |
|---|
Frühentwicklung
BearbeitenBildung der zweiblättrigen Keimscheibe
BearbeitenDie Zellen des Embryoblasten differenzieren sich zu zwei funktionellen Einheiten:
- die Endometrium-nahen Zellen bilden die eine Einheit: primitives Ektoderm (= Epiblast)
- die Endometrium-fernen Zellen bilden die andere Einheit: primitives Entoderm (= Hypoblast)
Beide Einheiten werden zur "Keimscheibe" zusammengefasst. Mit Ausbildung der Keimscheibe ist auch die ventro-dorsale Achse des Embryos erkennbar.
Nun bilden sich zwei Höhlen: erstens der primäre Dottersack, bei dem es sich um die ausgekleidete Blastocystenhöhle handelt, zweitens die Amnionhöhle zwischen Epiblast und Cytotrophoblast.
- Zellen des Hypoblasten breiten sich aus und kleiden dadurch die Blastocystenhöhle als "Dottersackepithel" oder "Heuser-Membran" von innen aus; die Blastocystenhöhle wird fortan als "primärer Dottersack" bezeichnet (nur vorübergehend vorhanden)
- Auf der anderen Seite entsteht ein kleiner flüssigkeitsgefüllter Hohlraum, der den Epiblasten vom Cytotrophoblasten trennt (primäre Amnionhöhle). Anschließend wandern Zellen des Epiblasten aus und kleiden diese primäre Amnionhöhle von innen aus, wodurch die primäre Amnionhöhle zur sekundären Amnionhöhle wird. Die sie auskleidenden Zellen nennen sich "Amniocyten" (in der Gesamtheit "Amnionepithel" oder "Amnioblast") und produzieren Fruchtwasser (am Ende der Schwangerschaft: 1 Liter; eine Amniozentese (Fruchtwasseruntersuchung) ist etwa ab der 15. Schwangerschaftswoche möglich).

|
|
Merke Indem sich die Zellen des Embryoblasten in Epi- und Hypoblast differenzieren, entsteht die zweiblättrige Keimscheibe. Der Hypoblast wird durch den primären Dottersack vom Trophoblasten getrennt, der Epiblast durch die Amnionhöhle. Beide Hohlräume sind ausgekleidet mit Zellen, die der jeweiligen Keimschicht entstammen. |
|---|
Bildung des extraembryonalen Mesoderms
Bearbeiten
Zellen des Hypoblasten wandern am 13. Tag aus und lagern sich zwischen Cytotrophoblast und Heuser-Membran/Amnionepithel als extraembryonales Mesoderm ein; wenig später bekommt es Löcher, die daraufhin zu einem Hohlraum zusammenfließen ("extraembryonales Zölom"). Das extraembryonale Mesoderm ist folglich in zwei Blätter gespalten, und zwar in
- das viszerale Blatt, das der Heuser-Membran und der Amnionepithel außen aufliegt, und
- das parietale Blatt, das dem Cytotrophoblasten innen anliegt.
Zwischen beiden Blättern bleibt nur eine einzige Verbindung bestehen, und zwar der Haftstiel, aus dem in der 4. Woche mitsamt eingelagertem Inhalt die Nabelschnur hervorgehen wird. Ein Teil der Heuser-Membran stülpt sich in den Haftstiel vor und wird als "Allantois" bezeichnet; diese bildet sich mit der Zeit zurück, am ausgewachsenen Menschen erinnert nur mehr das obliterierte Ligamentum umbilicale medianum an sie (pathologisch: Urachus-Fistel mit Harnaustritt aus dem Nabel).
Ab jetzt unterscheidet man zwei periphere Bereiche:
- Die mit Flüssigkeit gefüllte Chorionhöhle (ehemaliges extraembryonales Cölom, bleibt bis zum 3. Monat bestehen) zwischen beiden Blättern des extraembryonalen Mesoderms; mit dem Wachstum des Keimes dehnt sie sich stark aus und schafft somit gewissermaßen Platz für das Wachstum des Embryos.
- Das Chorion, das sich an die Chorionhöhle außen anschließt und aus den peripheren Bestandteilen des Keims besteht:
- parietales Blatt des extraembryonalen Mesoderms sowie
- Cyto- und Syncytiotrophoblast (eine Chorionbiopsie ist ab der 9.-12. Woche möglich).
Das Chorion bildet somit die Wand des Keimes und ist die Grundlage für den fetalen Teil der Plazenta. Als "Chorionzotten" bezeichnet man diejenigen Teile des parietalen extraembryonalen Mesoderms, die sich in Ausstülpungen des Cytotrophoblasten befinden.
|
|
Merke Das extraembryonale Mesoderm trennt zunächst die Einheit aus Keimscheibe, primärem Dottersack und Amnionhöhle vom Trophoblast ab. Später wird es durch die Chorionhöhle in ein viszerales und ein parietales Blatt getrennt, wobei das parietale Blatt zusammen mit den beiden Anteilen des Trophoblasten die äußere Wand (Chorion) des Keimes bildet. |
|---|
Entstehung der dreiblättrigen Keimscheibe (Gastrulation)
Bearbeiten
In der 3. Woche wandern Zellen des Epiblasten mittig zwischen Epiblast und Hypoblast und bilden dadurch das Mesoderm. Aus diesem gehen später u. a. die inneren Organe hervor.
In der Aufsicht auf den Epiblasten ist die Einwanderungsstelle zu sehen: Zunächst verdichten sich median gelegene Zellen des Epiblasten zum Primitivstreifen, der im Primitivknoten endet – somit ist die kranio-kaudale Achse des Embryos erkennbar. Dann senken sich die Zellen des Primitivstreifens und -knotens ein, es entstehen somit die Primitivrinne (teilt den Primitivstreifen in zwei Wülste) und die Primitivgrube (im Primitivknoten). Die sich einsenkenden Zellen haben zwei Ziele:
- Die Zellen aus dem Primitivstreifen wandern zwischen Epiblast und Hypoblast und werden zu intraembryonalem oder extraembryonalem Mesoderm (s. o.); das intraembryonale Mesoderm trennt Epi- und Hypoblast voneinander und fehlt nur kranial (Buccopharyngealmembran) und kaudal (Kloakenmembran), d. h. an diesen Stellen grenzen Epi- und Hypoblast direkt aneinander. Die verschiedenen Anteile des Mesoderms lassen sich bestimmten Abschnitten des Primitivstreifens zuordnen:
- kraniales Drittel des Primitivstreifens - paraxiales Mesoderm
- kranialer Anteil des mittleren Primitivstreifen-Drittels - intermediäres Mesoderm
- kaudaler Anteil des mittleren Primitivstreifen-Drittels - Seitenplattenmesoderm
- kaudales Drittel des Primitivstreifens - Anteile des extraembryonalen Mesoderms
- Die Zellen aus dem Primitivknoten dringen unterhalb des Primitivknotens in den Hypoblasten ein (indem sie die Hypoblast-Zellen zur Seite drängen) und wandern zunächst gewissermaßen "im Hypoblasten" nach kranial. Später wandert der Primitivknoten nach kaudal, wobei sich die vom Primitivknoten auswandernden Zellen parallel zur Wanderungsrichtung des Primitivknotens in die Zellschicht des Hypoblasten einlagern. Dadurch entsteht letztlich eine schmale Platte aus Epiblastzellen, die den Hypoblasten in kranio-kaudaler Richtung durchzieht. Die seitlichen und medianen Anteile dieser Platte haben ein unterschiedliches Schicksal:
- Die seitlichen Zellen proliferieren stark, verdrängen dabei den Hypoblasten und bilden schließlich das Entoderm.
- Aus dem medianen Anteil (Chordaplatte, Chordafortsatz) geht die Chorda dorsalis hervor; die Chordaplatte schnürt sich dabei als Rohr ab, wobei sich die während dieser Abschnürung entstandene Entoderm-Lücke schließt. Dieses als "Chorda dorsalis" bezeichnete Rohr liegt nun mittig im Mesoderm. Es dient als primäre Körperachse und induziert benachbarte Bereiche zu Differenzierungen (z. B. im Rahmen der Neurulation, s. u.). Die Chorda dorsalis geht im weiteren Verlauf fast vollständig zugrunde, im vollendeten Organismus erinnert nur mehr der Nucleus pulposus der Zwischenwirbelscheiben an sie.
Vorübergehend existiert zwischen Primitivgrube und kranialeren Abschnitten des Hypoblasten ein Kanal (Canalis neurentericus), so dass Amnionhöhle und Dottersack kurzzeitig miteinander in Verbindung stehen.
|
|
Merke Primitivstreifen und sein kranialer Endpunkt, der Primitivknoten, sind mesenchymale Verdichtungen des Epiblasten. Aus beiden Objekten gehen Zellen des Meso- und Entoderms hervor: Zellen des Primitivstreifens wandern zwischen Epi- und Hypoblast und bilden das Mesoderm, wobei mediale Mesoderm-Anteile von kranialen Primitivstreifen-Abschnitten abstammen und laterale Mesoderm-Anteile Derivate kaudalerer Primitivstreifen-Abschnitte sind; Zellen des Primitivknotens wandern in den Hypoblasten ein und bilden sowohl das Entoderm als auch den Chordafortsatz, der sich später als Chorda dorsalis abschnürt, eine Struktur, die in erster Linie Nachbarregionen zur Differenzierung induziert und später bis auf die Nuclei pulposi zugrunde geht. |
|---|
Die folgenden Vorgänge vollziehen sich etwa zur gleichen Zeit.
Differenzierung des intraembryonalen Mesoderms
BearbeitenDas Mesoderm gliedert sich, ausgehend von der Chorda dorsalis, von medial nach lateral in
- das paraxiale Mesoderm,
- das intermediäre Mesoderm und
- das Seitenplattenmesoderm.
Jeder dieser drei Teile formt sich auf charakteristische Weise um.
Differenzierung des paraxialen Mesoderms
BearbeitenNach Induktion durch die Chorda dorsalis formt sich das paraxiale Mesoderm von kranial nach kaudal zu Somiten um, d. h. zu miteinander verbundenen würfelförmigen, hohlen Epithelstrukturen, denen sich außen eine Basalmembran auflagert. Dadurch ist die Metamerie des Körpers festgelegt.
Während nach kaudal hin weitere Somiten entstehen, werden die kranialen Somiten von umgebenden Strukturen dazu induziert, sich weiter zu differenzieren, und zwar in ein ventromediales Segment (Sklerotom) und ein dorsolaterales Segment (Dermomyotom).
- Das Sklerotom entdifferenziert sich zu Mesenchymzellen, wandert dann nach medial und vereinigt sich mit den Zellen des kontralateralen Sklerotoms, um später Rippen und Wirbel zu bilden.
- Das Dermomyotom differenziert sich zu zwei Anteilen, und zwar
- zum nach außen hin gelegenen Dermatom, aus dem später die Dermis hervorgeht, und
- zum nach innen hin gelegenen Myotom, aus dem sich Muskeln entwickeln, und zwar einerseits autochthone Muskeln (Epimer) und andererseits eingewanderte Rumpf- und Extremitätenmuskeln (Hypomer).
Differenzierung des intermediären Mesoderms
BearbeitenDas intermediäre Mesoderm (Somitenstiel) differenziert sich zu den Vorläufern der Urogenital-Organe:
- der kraniale Teil differenziert sich zu segmental geordneten Nephrotomen,
- der kaudale Teil bildet einen unsegmentierten Strang, den Urnierengang; hierbei handelt es sich um ein Blastem, d. h. um eine Mesenchymverdichtung, die sich später differenzieren, ausdehnen und umformen kann.
Später entstehen hieraus Vorniere, Urniere und Nachniere sowie die Ureteren.
Differenzierung des Seitenplattenmesoderms
BearbeitenIm Seitenplattenmesoderm ist zunächst eine geschlossene Zellmasse, das den Anschluss an das extraembryonale Mesoderm gewährleistet. Später entsteht im Seitenplattenmesoderm ein Spalt (intraembryonales Zölom), welcher das Seitenplattenmesoderm in zwei Blätter unterteilt:
- ein parietales Blatt (Somatopleura), das dem Ektoderm aufliegt; es bildet den Vorläufer für das Bindegewebe der vorderen Rumpfwand sowie für das Sternum.
- ein viszerales Blatt (Splanchnopleura), das dem Entoderm aufliegt; aus ihm gehen später das Bindegewebe und die Muskulatur des Gastrointestinaltrakts hervor, zudem bilden sich hier die Blutinseln.
Kaudal sind beide Blätter vollständig voneinander getrennt, so dass intraembryonales und extraembryonales Zölom (Chorionhöhle) direkt miteinander in Verbindung stehen. Kranial ist die Trennung unvollständig, denn zwischen beiden Blättern existiert das Septum transversum.
Abfaltung der Keimscheibe
Bearbeiten-
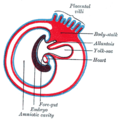 Kraniokaudale Abfaltung
Kraniokaudale Abfaltung -
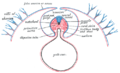 Laterale Abfaltung
Laterale Abfaltung -
 Ergebnis der Abfaltungsprozesse
Ergebnis der Abfaltungsprozesse



In der vierten Embryonalwoche faltet sich die dreiblättrige Keimscheibe sowohl in kranio-kaudaler als auch in lateraler Richtung ab, wodurch aus Anteilen des primären Dottersacks das Darmrohr hervorgeht.
Kraniokaudale Abfaltung
BearbeitenDas kraniale (Kopffalte) und das kaudale Ende (Schwanzfalte) der Keimscheibe stülpen sich zunächst über die mittleren Anteile und nähern sich dann auf der Ventralseite einander an, ohne miteinander zu verschmelzen. Dadurch wird der Dottersack zwar nicht abgeschnürt, aber dennoch stark eingeschnürt, so dass sein Entoderm-naher Teil eine Röhre bildet. Dies ist das Darmrohr, das sich in drei Abschnitte gliedert, und zwar in
- einen Vorderdarm,
- einen Mitteldarm (Übergang zwischen Vorderdarm und Mitteldarm: vordere Darmpforte) und
- einen Hinterdarm (Übergang zwischen Mitteldarm und Hinterdarm: hintere Darmpforte).
Da der Dottersack nicht vollständig abgeschnürt wird, bleibt zwischen Mitteldarm und dem restlichen Dottersack eine Verbindung bestehen (Ductus omphaloentericus, Ductus vitellinus), die jedoch später obliteriert.
Am kranialen Ende des Vorderdarms sowie am kaudalen Ende des Hinterdarms fehlt Mesoderm, weshalb Ekto- und Entoderm hier direkt aneinander grenzen und somit die Bukkopharyngealmembran (Rachenmembran, kranial) sowie die Kloakenmembran (kaudal) bilden.
- Die Bukkopharyngealmembran wird zunächst durch die stark wachsenden Gehirnbläschen in die Tiefe verdrängt. Gleichzeitig bildet sich vor ihr durch Auswachsen der Kiefer- und Gesichtsfotsätze das Stomatodeum (Mundbucht, Vorläufer von Mundhöhle und Teilen der Nasenhöhlen). Anschließend reißt die Bukkopharyngealmembran ein, so dass sich Stomatodeum und Vorderdarm miteinander verbinden.
- Hinter der Kloakenmembran bildet sich ebenfalls eine Einsenkung, und zwar das Proktodeum (Afterbucht). Auch die Kloakenmembran reißt ein, so dass sich Proktodeum und Hinterdarm miteinander verbinden. Die Kloakenmembran wird zusätzlich durch das von kranial einwachsende Septum urorectale in einen ventralen Abschnitt (Vorläufer von Harnblase und Harnröhre) und einen dorsalen Abschnitt (Vorläufer des Anorektalkanals) getrennt.
Laterale Abfaltung
BearbeitenDie laterale Abfaltung vollzieht sich sowohl in peripheren Bereichen des Embryos, wodurch die Körperwand festgelegt wird, als auch in zentralen Bereichen, wodurch das Darmrohr festgelegt wird. An den entsprechenden Abfaltungsvorgängen sind entweder Ekto- oder Entoderm entscheidend beteiligt.
- Die seitlichen Anteile des Ektoderms sowie die an ihm anliegenden Teile des intraembryonalen Mesoderms (parietales Blatt des intraembryonalen Mesoderms) wandern zunächst seitlich gewissermaßen um das Mesoderm herum, nähern sich dann auf der Ventralseite dem Dottersack, der durch eine analoge Abfaltung des Entoderms abgeschnürt wird (s. u.), und verschmelzen schließlich miteinander. Somit umgibt das Ektoderm sowohl das Mesoderm als auch das Darmrohr, wobei das Ektoderm seinerseits vollständig von der Amnionhöhle umgeben wird.
- Gleichzeitig zum gerade beschriebenen Vorgang wandern die seitlichen Anteile des Entoderms sowie die ihm anhaftenden Teile des intraembryonalen Mesoderms (viszerales Blatt des intraembryonalen Mesoderms) um den Dottersack herum, schnüren ihn zu großen Teilen ab (Ausnahme: Ductus omphaloentericus, s. o.) und umhüllen ihn.
Neurulation
Bearbeiten

Die Chorda dorsalis induziert die Zellen des Epiblasten. Diese
- differenzieren sich zu Neuroepithel (Neuralplatte),
- heben sich in Form von zwei Falten aus der Epiblast-Ebene ab (Neuralfalten), wobei sie eine Rinne begrenzen (Neuralrinne),
- die Seiten der Neuralfalten wandern aufeinander zu und schließen sich zum Neuralrohr; die Zellen des Epiblasten verbinden sich anschließend wieder, so dass das Neuralrohr nun als eigenständiges Objekt unterhalb der Epiblasten-"Decke" im Mesoderm vorliegt. Teile des Neuralrohrs spalten sich als Neuralleiste ab.
Der Verschluss des Neuralrohrs erfolgt nicht an allen Stellen gleichzeitig, sondern vielmehr reißverschlussartig: das Rohr wird von der Mitte aus (4. Somit) nach kranial und kaudal "zugezogen", in der Folge spaltet sich dann auch die Neuralleiste ab. Kranial und kaudal bleiben dabei zunächst zwei Öffnungen bestehen, die sich später verschließen (Neuroporus cranialis: Verschluss am 24. Tag; Neuroporus caudalis: Verschluss am 26. Tag).

Das Neuralrohr ist die Grundlage des zentralen Nervensystems:
- Aus dem kaudalen Anteil entwickelt sich das Rückenmark
- Aus dem kranialen Anteil bildet sich das Gehirn; zunächst liegen drei Gehirnbläschen vor, die sich dann weiter aufteilen (von kaudal nach kranial: Rhombencephalon (wird später zu Medulla oblongata und Metencephalon), Mesencephalon, Prosencephalon (später Diencephalon und Telencephalon)). Die Gehrinbläschen wachsen dabei so stark, dass sie die Herzanlage vollständig bedecken.
Die Derivate der Neuralleiste sind demgegenüber sehr heterogen:
- Periphere Neurone
- Neurone der Spinalganglien
- Neurone des vegetativen Nervensystems, auch Zellen des Nebennierenmarks (sind funktionell postganglionäre Neurone)
- Neurone der Hirnnerven-Ganglien
- Gliazellen
- Schwannzellen (wickeln sich als Myelinscheide um periphere markhaltige Neurone)
- Mantelzellen (umgeben die Perikaryen der Spinalganglien)
- Kopfmesenchym (Hirnhäute, Dermis, Knochen)
- Melanocyten (Haut)
- C-Zellen (Schilddrüse)
- Odontoblasten (Zahn)